1. fejezet - A háromdimenziós technikák pszichológiai és fiziológiai háttere
- 1.1. Az emberi érzékelés
- 1.2. Az emberi észlelés
- 1.3. Az emberi szem felépítése és működése
- 1.4. A látópálya felépítése és működése
- 1.5. A színlátás
- 1.6. A háromdimenziós mélységészlelés
- 1.7. Az emberi mélységészlelés hibái
- 1.8. Összefoglalás
- 1.9. A felkészülést segítő kérdések
- 1.10. A fejezet fontosabb kifejezései
- Felhasznált irodalmak
Mivel ez a könyv elsősorban a műszaki érdeklődésű fiataloknak íródott, fontosnak éreztük, hogy mielőtt belevágunk a mérnök hallgatóknak már megszokott technológiai fogalmak ecsetelésébe, adjunk egy átfogó bevezetést a releváns pszichológiai és fiziológiai folyamatokba, amelyek nélkülözhetetlenek a háromdimenziós élmény létrejöttéhez. Ezzel reméljük, csak közelebb visszük az Olvasót a teljes folyamat megértéséhez és felkeltjük érdeklődését az interdiszciplináris tudományok iránt.
Jelen fejezetben elsősorban azokra a nem műszaki területekre koncentrálunk, amelyek hozzájárulnak a háromdimenziós élmény létrejöttéhez. Részletezzük az emberi szem felépítését és működését, amelyen keresztül az információ megérkezik a külvilágból elektromágneses sugárzás formájában. Megismerkedünk azon agyi folyamatokkal, idegpályákkal, amelyen keresztül agyunk feldolgozza a látottakat és lehetővé teszi az információ tudatosodását. Mivel számos technika alapul a színszűrés elvén, megismerkedünk az emberi színlátás mechanizmusával. Végül a legfontosabb rész következik az emberi mélységészlelés folyamatáról, amely lehetővé teszi, hogy egyáltalán háromdimenziós élményben legyen részünk.
A pszichológián kívül sok tudományterület foglalkozik a tudatunkkal, viselkedésünkkel. Ilyenek például a biológia, a nyelvészet, a filozófia, az orvostudományok és napjainkban egyre szélesebb körben a különböző mérnöki tudományok. Ez a kapcsolódás nem csupán a tervezett eszközök, készülékek kényelmi szempontjait érinti, hanem a teljes ember-gép kapcsolatrendszert, amelynek szerves része többek között az ember és intelligens berendezései közötti kommunikáció hatékony megvalósítása. Napjainkban egyre több olyan műszaki kutatás, mérnöki fejlesztés létezik, amelynek lételeme a pszichológia – etológia, viselkedéstudomány és a kognitív tudományok. Csak két kiemelkedő példát említve: etorobotika – etológia és mechatronika – és kognitív informatika – pszichológia és informatika. Ezért a „jövő mérnökeinek” elengedhetetlen az interdiszciplináris gondolkodás mielőbbi elsajátítása.
Jelen fejezetben beszélni fogunk az emberi érzékelés és észlelés különbségéről, az emberi szem felépítéséről és működéséről, a látópálya felépítéséről és működéséről, a színlátásról, a háromdimenziós mélységészlelésről és végül az emberi mélységészlelés hibáiról.
1.1. Az emberi érzékelés
Az emberi érzékelés az a folyamat, amely során a külvilágból érkező információk – legyenek azok szagmolekulák, szöggyorsulás értékek, nyomás a bőrön vagy éppen elektromágneses sugárzás, fény – valamely érzékszervünkön keresztül – valamely modalitásban – elérhetővé válnak a feldolgozórendszerünk számára. Tehát a külvilág fizikai jelei alakulnak át a feldolgozó rendszerünk – központi idegrendszerünk – által befogadható jelekké. Ezt nevezzünk szignál transzdukciónak, vagyis szenzoros kódolásnak. Az érzékelés egy biológiai alapfolyamat, amely csupán az érzékszerveink által végzett folyamatokat érinti. Az érzékszerveink transzdukciót végző egységei a receptorok, amelyeknek modalitásonként eltérő a felépítésük és a működésük. A receptorok a fizikai jelekből csak a számukra megfelelő – adekvát – ingereket dolgozzák fel. A feldolgozás után már ingerületről beszélünk. Az ingerületek valójában neurális impulzusok, amelyek a központi idegrendszerbe továbbítódnak akciós potenciálok formájában.
1.2. Az emberi észlelés
Az észlelési folyamat – percepció – magából az érzékelésből és a hozzá kapcsolódó kognitív feldolgozásból áll. Amikor nézünk valamit, még nem biztos, hogy látjuk is. Érzékelés történhet észlelés nélkül is, de ez fordítva nem igaz. Az észlelés során a különböző érzékleti mintázatokhoz rendeljük hozzá a külvilág eseményeit. Ha például egy meghatározott spektrumú hangot hallunk, tudjuk, hogy valószínűleg mit hallunk. Az észlelés tehát egy pszichológiai alapfolyamat, amelynek az érzékelés az előfeltétele.
1.3. Az emberi szem felépítése és működése
Ebben az alfejezetben az emberi szemről és annak számos tulajdonságáról olvashatunk. Szó lesz az érzékelhető elektromágneses sugárzásról, az emberi szem elhelyezkedéséről és a speciális szemmozgásokról, az emberi szem felépítéséről és anatómiájáról, továbbá a retináról, a fotoreceptorokról és végül a látási hibákról.
1.3.1. Az érzékelt elektromágneses sugárzás
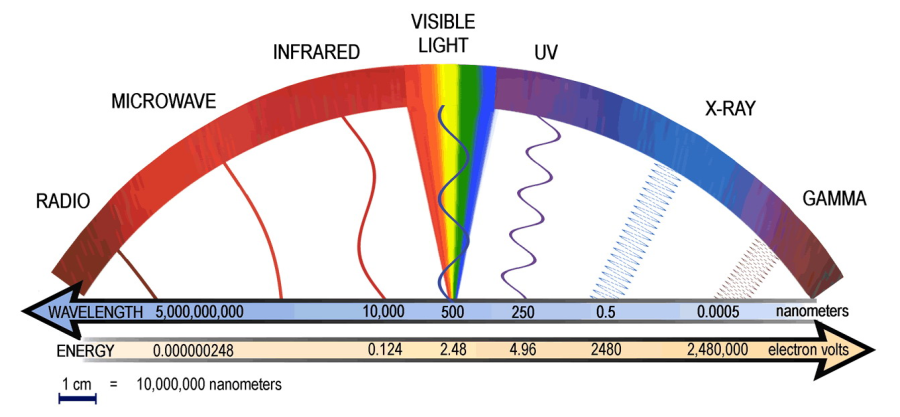
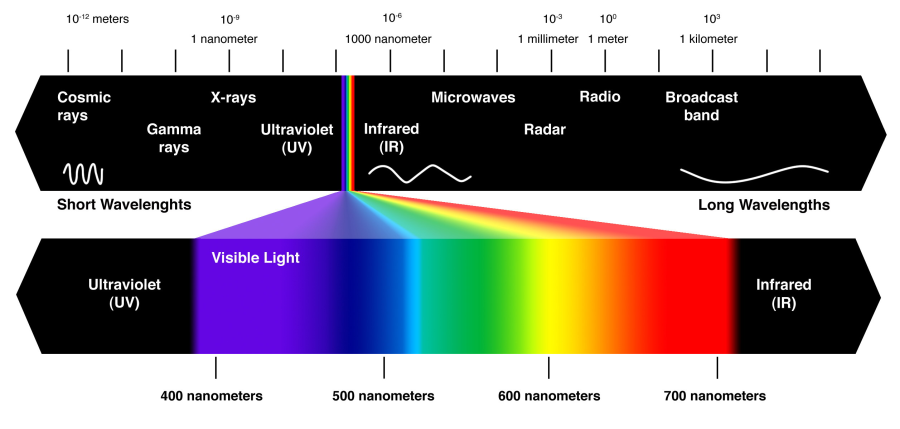
Az emberi szem a teljes elektromágneses spektrumból hozzávetőlegesen csupán a 380 és 780 nm közötti hullámhossz-tartományt képes érzékelni. Ez a látható színtartomány az ultraibolyától, az infravörösig terjed. A Földön honos élőlények nagy része ebben a tartományban képes érzékelni a környezetéből érkező sugárzást. Ez nagy valószínűséggel annak köszönhető, hogy a 400 nm alatti sugárzást jó részét a légkör elnyeli, a 800 nm feletti sugárzás nagy része pedig áthatol a tárgyakon. Az általunk látható fény általában összetett fény, amely több, különböző frekvenciájú sugárzást tartalmaz. Ilyen fényt sugároz magából a Napunk is, amelynek spektrumára a törzsfejlődésünk során adaptálódott a látórendszerünk. Amennyiben a fény csak egy hullámhosszú, azt monokromatikus fénynek nevezzük. Léteznek továbbá úgynevezett kvázi monokromatikus sugárzások is, amelyek néhány nanométeres intervallumban tartalmaznak különböző frekvenciájú összetevőket. A nagy frekvenciájú fényhez kisebb hullámhossz érték, míg a kis frekvenciájúhoz nagyobb hullámhossz-érték tartozik.
1.3.2. Az emberi szem elhelyezkedése és a szemmozgások
Az állatvilágban nagyon változatos látószerveket ismerhetünk meg, amelyek elhelyezkedése és felépítése nagymértékben eltér egymástól. Amíg a növényevő állatok szemei úgy helyezkednek el a fejen, hogy a lehető legnagyobb belátható teret legyenek képesek megfigyelni, addig a ragadozók szemei általában a fejük elülső részén helyezkednek el, egymáshoz közelebb, hogy képesek legyenek megbecsülni prédájuk távolságát a sikeresebb vadászat érdekében, ehhez ugyanis három dimenzióban való látásra van szükség.
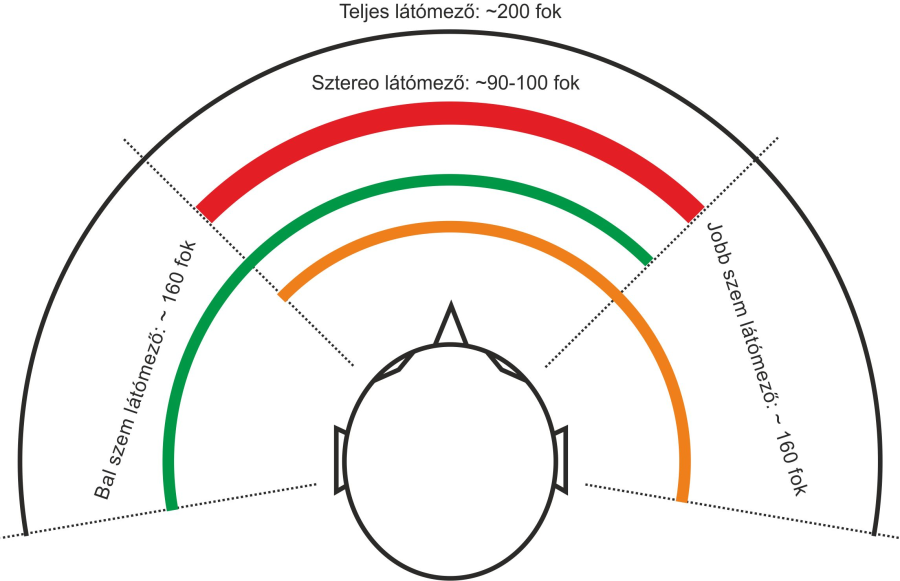
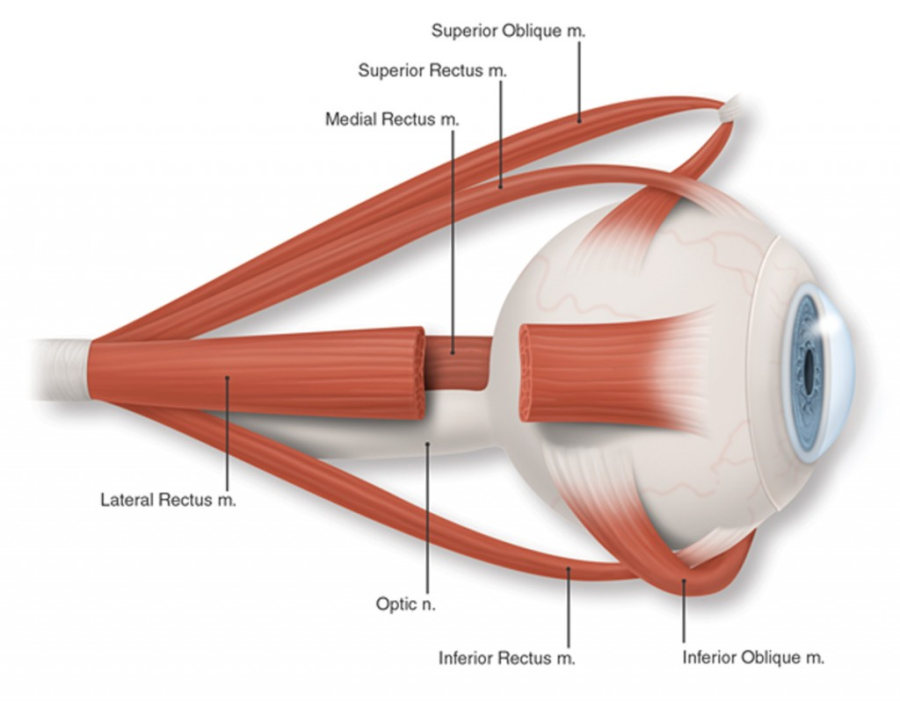
Az emberi szem a fej elülső oldalán helyezkedik el, egymással egy síkban, és egymástól átlagosan 63 mm-es távolságban. Ezzel a látómezőnk közel 90 fokban, sztereo látást tesz lehetővé, tehát eme tartományban mindkét szemünkkel látjuk a tárgyakat, ezen kívül legfeljebb csak az egyik szemünkkel – monokuláris látómező 160 fok, binokuláris látómező 200 fok. Szemünk mozgását a szem körül elhelyezkedő – szemenként – három pár extraokuláris izom teszi lehetővé. Ezek a mozgások jellemzően nem folyamatos, pásztázó mozgások, hanem szakaszosak. Létezik egyirányú – konjunktív – szemmozgás, amely során a két szemünk egy irányba mozog, és létezik ellentétes irányú – vergens – szemmozgás, amely során szemünk a közeledő, illetve távolodó tárgyakra néz. A közvetlenül előttünk lévő tárgya történő fókuszálás a konvergens szemmozgás.
Szemünk két-féle szemmozgást képes kivitelezni. Az egyik egy szakaszos, szaggatott, gyors szemmozgás, amely például olvasáskor vagy a weboldalakon való böngészéskor aktív. A másik egy folyamatos mozgás, amely a mozgó tárgyak követéséhez nélkülözhetetlen. A szakaszos szemmozgás ugrásait szakkádoknak, a megállásait fixációnak nevezzük. A szakkádikus elnyomás következtében a gyors szemmozgások alatt nincs információ-felvétel. A később bemutatott retinális mechanizmusoknál látni fogjuk, hogy a fotoreceptoroknak folyamatos „frissítésre” van szükségük ahhoz, hogy a fixált képet érzékeljék. Ehhez nélkülözhetetlen egyfajta mikroszakkádikus mozgás, amelynek következtében a szem soha nincs teljesen nyugalomban. Amennyiben a mikroszakkádokat kiiktatnánk az észlelt tárgyak elhalványulnának és előbb-utóbb el is tűnnének a látómezőből. A követő szemmozgáskor a mozgás sebessége és iránya folyamatosan változik.
1.3.3. Az emberi szem felépítése, anatómiája
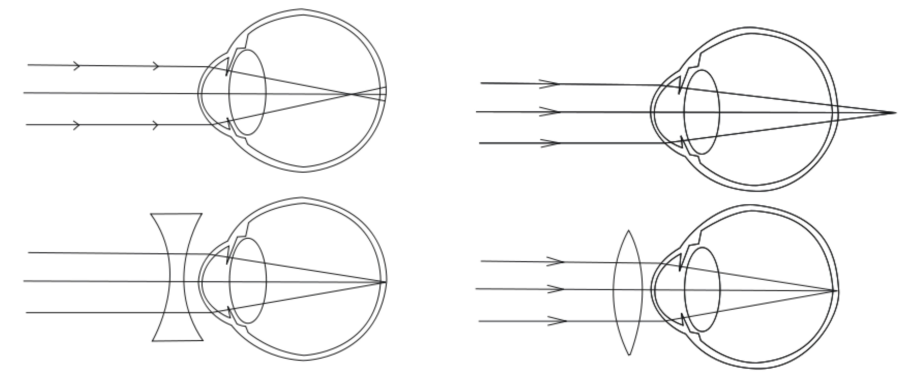
Az emberi szem anatómiáját tekintve végső soron három koncentrikus rétegből – ínhártya, érhártya, retina – két kamrából – elülső szemkamra, üvegtest – a szivárványhártyából, a pupillából és a lencséből áll. Az ínhártya a szem külső, rostos, kemény rétege. Ennek elülső része voltaképpen a szaruhártya – cornea – amelynek már nincs saját vérellátása, az elülső kamrai folyadék táplálja. Optikai szempontból a szaruhártya adja nagy részét a szem dioptriaértékének – kisebb részt ad a szemlencse. Az érhártya a szemgolyó falának közelében tapad, hajszálerei táplálják az ínhártyát. Az elülső szemkamrában található a csarnokvíz, amely a cornea táplálása a szem alakjának fenntartásában is rész vállal – az ínhártya mellett. Az üvegtest a szem tömegének közel egy-harmada. A benne lévő folyadék szintén csarnokvíz, de azzal ellentétben nem újul meg folyamatosan. A szivárványhártya a szemlencse elülső oldalára simul rá és ez adja szemünk jellegzetes színét. A közepén lévő fekete terület a pupilla, amely valójában két izomcsoport közötti kerek rés, nyílás. A pupilla mérete befolyásolja, hogy mennyi fény éri el a retinát. A szemlencse a szivárványhártya mögött helyezkedik el. Átlagosan 9 mm átmérőjű és 4 mm vastag. A szem a szemlencse segítségével képes a fokuszálásra, amelynek következtében élesen láthatunk. Ezt az élesre állítási folyamatot hívjuk akkomodációnak. A lencse alakját, illetve görbületi sugarát a ciliáris izmok segítségével változtatjuk meg. A szemlencsét a lencsefüggesztő rostok tartják a helyén megfeszített állapotban. Közelre nézéskor a ciliáris izmok megfeszülnek, és a lencsefeszítő rostok elernyednek, melynek következtében a szemlencse magától összezsugorodik, ezzel csökkentve a görbületi sugarat. Távolra nézéskor egy ellentétes folyamat játszódik le: a ciliáris izmok elernyednek, a lencsefeszítő rostok megfeszülnek, melynek következtében a szemlencse kinyúlik, ezzel növelve a görbületi sugarat. Ezért van az is, hogy ha elfáradunk a ciliáris izmok nem képesek a feszítésre, így csak „bámulunk a semmibe”, a fókuszunk a távoli célponton marad.
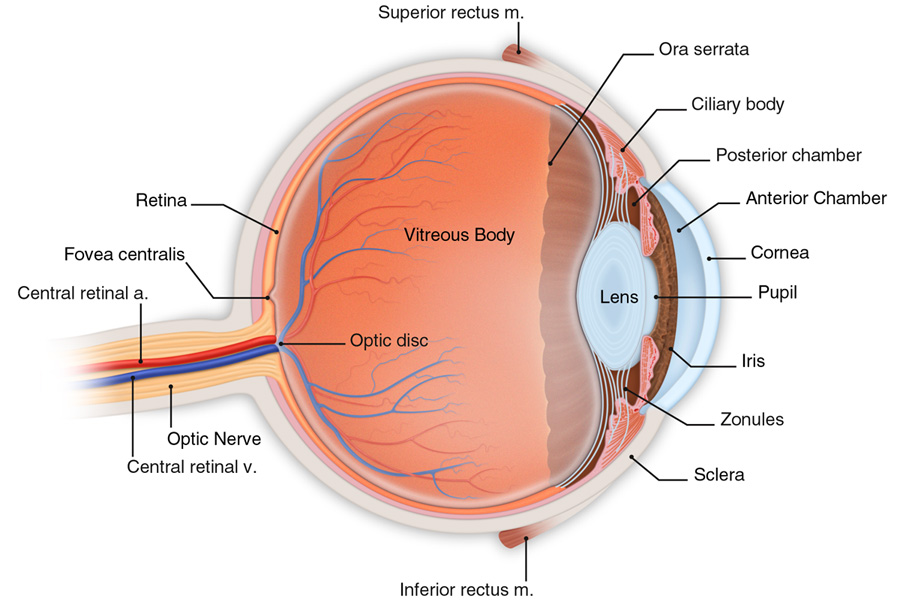
1.3.4. A retina és a fotoreceptorok
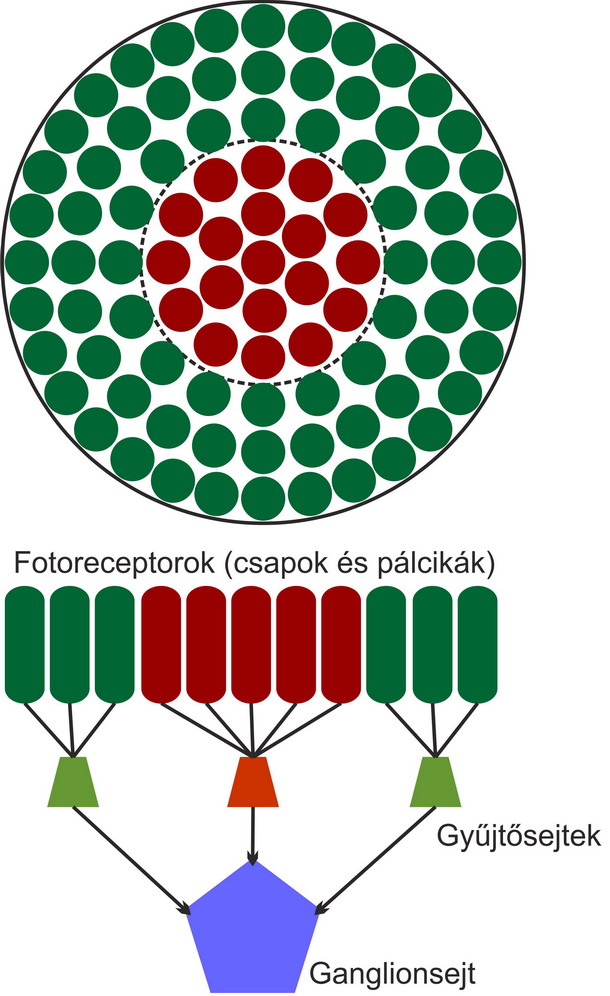
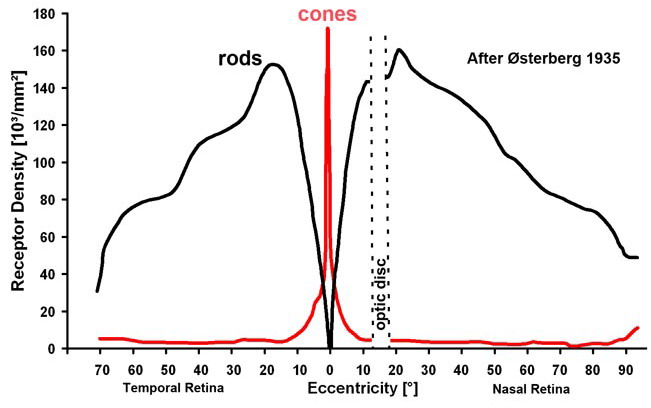
A látás receptorai a fotoreceptorok a retinán helyezkednek el. Ezek a receptorok a fényenergiát idegi jelekké alakítják át, majd továbbítják a közelben elhelyezkedő gyűjtősejtekből álló hálózatba. A retinán lévő fotoreceptorokhoz a fény a retinális ganglionsejteken és a gyűjtősejteken – bipoláris, amakrin és horizontális sejtek – keresztül érkezik. A retinán az éleslátás helye az úgynevezett sárgafolt vagy macula – fovea. Ahol a gyűjtősejtek idegrostjai elhagyják a retinát – vakfolt – az agy felé nincsenek fotoreceptorok.
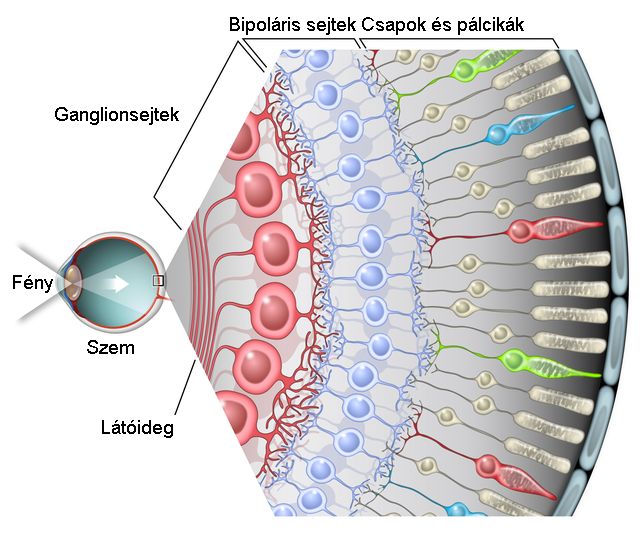
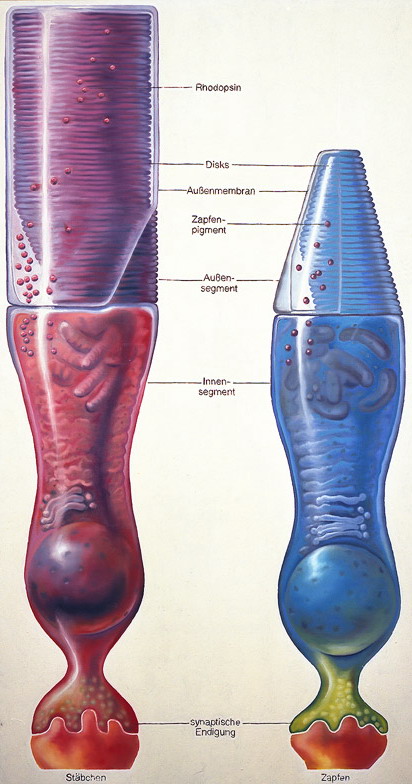
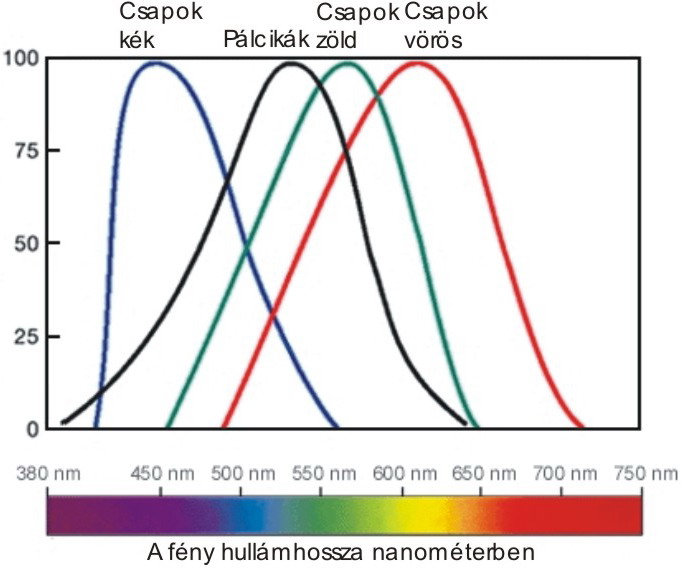
A retinán lévő fotoreceptorok száma hozzávetőlegesen 130 millió – 120 millió pálcika és 10 millió csap. A receptorok két fő típusa a csapok és pálcikák. A pálcikák több fotont képesen elnyelni több irányból, egy ganglionsejthez több pálcika idegrostja fut be, ezért erősítésük is jobb. Elsősorban a pálcikák felelősek a mozgás és fényesség érzékeléséért. A csapok három altípusát különböztetjük meg. Léteznek rövid, közép és hosszú hullámhosszakra érzékeny csapok, amelyek ezért a látható hullámhossztartományban elhelyezkedő kék, zöld és vörös színekkel azonosíthatóak. A zöld és vörös színekre érzékeny csapok érzékenységi tartományai igen közel vannak egymáshoz, ezért érthető, hogy miért alakulhat ki vörös-zöld színtévesztés. A kék színre érzékeny csapok jóval kevesebben vannak, ezért a kék fényben kevésbé látunk élesen. Az éleslátás helyén szinte csak csapok vannak, a periférián pedig szinte csak pálcikák. Ezért a fényintenzitás és mozgásészlelésünk a periférián, míg a színdiszkriminációs képességünk a foveán a legjobb. A fotoreceptorok sűrűsége a foveán a legnagyobb.
Ha a fotoreceptorokban lévő rodopszin – opszinból és retinalból áll – elnyel egy fotont, megváltozik a fehérje szerkezete és energia szabadul fel. Ezután a sejten belül elindít egy jelátviteli kaszkádot, amely számos fehérjét megváltoztat, majd kinyílnak a receptorsejt nátriumcsatornái és depolarizálódik. A neuronális jelet a fotoreceptor ezt követően a gyűjtősejtekhez továbbítja. A bipoláris, horizontális és amakrin gyűjtősejteken keresztül az ingerület a retinális ganglionsejtekhez továbbítódik – amelyekből 1 millió darab van a retinán – majd a vakfolton keresztül elhagyja a szemet az agy felé.
A tárgyaktól a retináig eljutó fényeloszlás voltaképpen a retinális kép. A tárgyak, amelyekről több foton érkezik fényesebbek, amelyekről kevesebb foton érkezik sötétebbek a látórendszer számára. A külvilág tárgyainak elemi pontjait reprezentáló fényintenzitás-eloszlást képnek hívjuk.
1.3.5. Látási hibák
Végezetül tegyünk említést a látási hibák két alaptípusáról! A normál látású, emmetropiás szem optikai rendszere a tárgyak képét pontosan a retinára képezi le. A nagyobb sugarú lencsével rendelkező, távollátó vagyhiperópiás szem a tárgy képét a retina mögé képezi le ezért a kép nem lesz éles, nem lesz fókuszált. Mivel ez esetben a szemlencse görbületi sugarát csökkenteni, konvexitását növelni kellene, a távollátó szem korrekciója konvex – domború – szemüveggel lehetséges. A rövidlátó, miópiás szem szemlencséjének görbületi sugara kisebb, ezért a tárgy képét a retina elé képezi le. Mivel ez esetben a lencse túlzottan is konvex, a korrekció konkáv lencsével korrigálható. Így a szemlencsét elhagyó sugarak kevésbé lesznek széttartóak, és a leképezés a retinára esik.
1.4. A látópálya felépítése és működése
Ebben az alfejezetben szót ejtünk a retinális ganglionsejtekről, a gyűjtősejtekről, a receptív mezőről, magáról a látópályáról és a látás további területeiről.
1.4.1. A retinális ganglionsejtek, a gyűjtősejtek és a receptív mező
Mint azt az imént kifejtettük, a fotoreceptorok neuronális jelei a gyűjtősejteken – bipoláris, horizontális és amakrin sejtek – keresztül a retinális ganglionsejtekhez jutnak. A ganglionsejtek száma körülbelül 1 millió. Mivel minden csap és pálcika jele eljut egy ganglionsejthez, ebből következik, hogy egy ganglionsejthez átlagosan 130 darab receptor jele továbbítódik a gyűjtősejtek közvetítésével. Stephen Kuffler és munkatársai – magyar származású orvos neurológus – 1952-ben fedezte fel azt a jelenséget, hogy a ganglionsejtekhez tartozó fotoreceptorok ingerlésekor a kisebb átmérőjű fényfoltra nagyobb reakciót adott a sejt. Ebből arra következtettek, hogy a ganglionsejtekhez kapcsolódó fotoreceptorok egy kvázi kör alakú, úgynevezett receptív mezőt alkotnak, amelynek a közepén lévő receptorokat ingerelve növelhető, a perifériát ingerelve pedig csökkenthető a ganglionsejt reakciója. Amennyiben az ingerlő fényfolt mérete akkora, hogy belelóg a szélső területekbe, a ganglionsejt gátlás alá kerül, és kisebb reakciót ad, mint ha csak a receptív mező közepét ingerelnénk. Ebből az következik, hogy a retina fotoreceptorai végső soron nem egyenként adnak visszajelzést a fényerősségről, hanem receptív mezőkbe tömörülve téri integrációt végeznek. A receptív mező elmélet egyik legismertebb demonstrációját az úgynevezett Hermann-rács adja. Később születtek elméletek, melyek szerint a Hermann-rács illúziója nem csupán a retinális feldolgozás eredménye, hanem magasabb agyi folyamatok is szerepet játszanak benne. A retinális kép feldolgozásának első és legjelentősebb állomása tehát a retinális ganglionsejtek lényeges változásokat kiemelő mozzanata. Kétféle retinális ganglionsejtet különböztetünk meg. Az egyik a receptív mező közepén ingerelve fokozza az aktivitását, a másik a receptív mező szélén ingerelve fokozza az aktivitását. Ezek az úgynevezett BE központú, KI szélű és KI központú, BE szélű ganglionok. A kétféle ganglionsejt teszi lehetővé többek között a világosságnövekedés és világosságcsökkenés egymástól független kódolását.
1.4.2. A látópálya
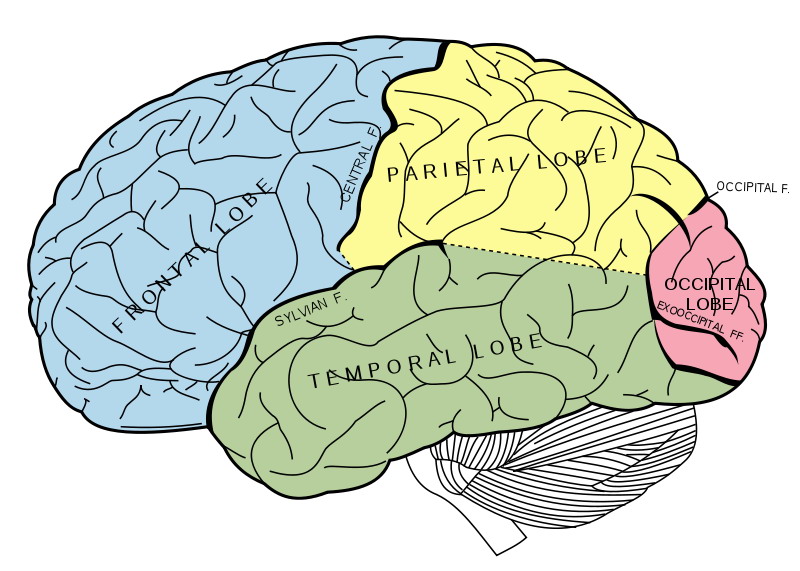
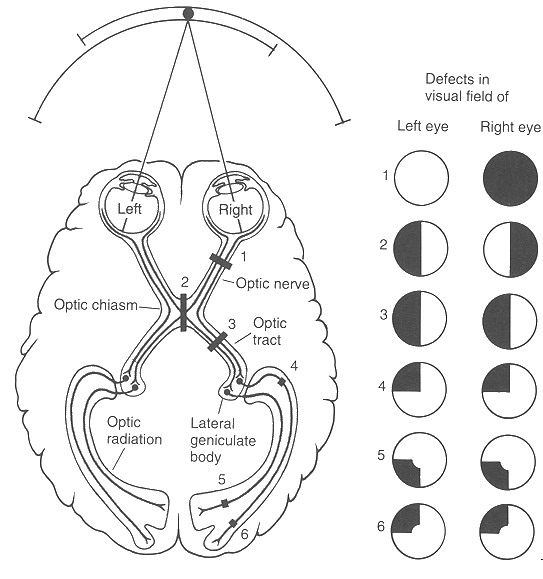
A retina és a látókéreg között a látópálya teremt kapcsolatot. Ez végső soron axonok – idegsejtek kommunikációs nyúlványai – kötege. A retinális ganglionsejtek axonjai alkotják tehát a látóideget. A látóidegek ez után kereszteződnek – bal és jobb szem idegkötegei – a caelsma opticumnál. Az orr felőli, nasalis részük ellentétes, a halánték felőli részük azonos oldalon halad tovább. Tehát tulajdonképpen a szembe balról érkező fény – kép – az agy jobb féltekéjébe tart. Innen egy részük az agytörzsbe megy, ahol a vegetatív folyamatok segítségére lesz, másik részük pedig a hipotalamuszban lévő CGL – corpus geniculatum laterale – területen átkapcsolódik. Az átkapcsolás után az idegrostok az úgynevezett látó kisugárzást alkotva a tarkólebenybe – okcipitális lebeny – tartanak, amelyet V1 területnek nevezünk.
1.4.3. A látás további területei
A V1 látórendszer egy úgynevezett retiotopikus reprezentációval rendelkezik, tehát a területet alkotó, egymás szomszédságában lévő neuroncsoportok a retinán is szomszédos receptorokból származnak. A V1 területen a kép további módosulása is bekövetkezik. Nevezetesen, az itt lévő neuroncsoportok képi reprezentációja során a foveáról származó információ túlreprezentálása következik be. Tehát az éleslátásunk területét „kinagyítja” a V1 terület. Ezt nevezzük agykérgi nagyításnak.
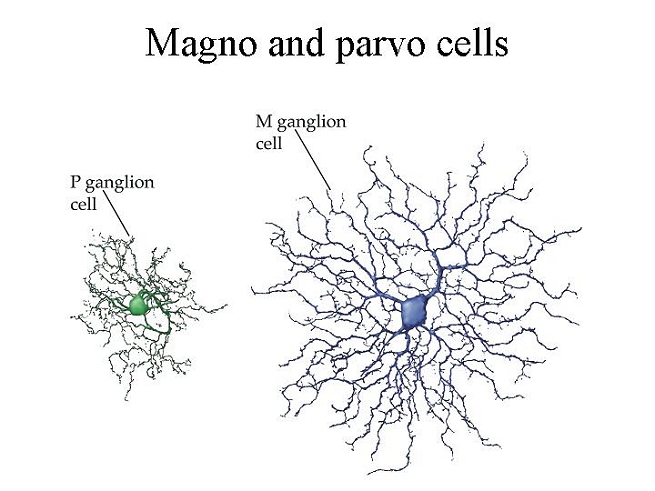
Egy másik felosztás szerint a ganglionsejteknek van egy kiterjedtebb dendritfával – gyűjtő neuronális nyúlványok – és egy kisebb dendritfával rendelkező típusa. A nagyobb receptív mezővel rendelkező ganglionsejteket M sejteknek, a kisebb receptív mezővel rendelkezőket P sejteknek hívjuk. Az M sejtek a CGL terület magnocelluláris rétegéhez, a P sejtek a CGL parvocelluláris rétegéhez kapcsolódnak. Mivel az M sejtekhez több receptor tartozik, azért a téri felbontóképességük rosszabb, mint a P sejteknek, viszont az M sejtek idői felbontóképessége sokkal jobb, mint a P sejteké, mert aktivitásuk kevesebb ideig marad fenn. Továbbá az M sejtek csak világosságot kódolnak, a P sejtek színeket is.
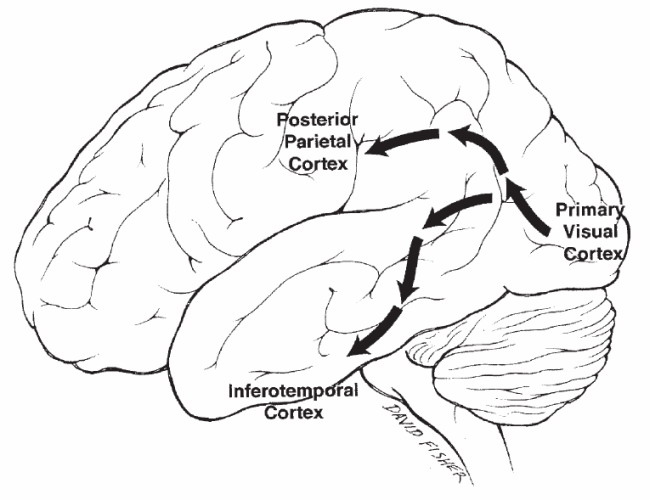
A V1 területről az információk agy része az egy felső részébe, a parientális lebenybe tart a dorzális területre, egy másik részük az agy alsó részébe, a temporális lebenybe tart a ventrális területre. A dorzális területre az M sejtek ingerületei tartanak, a ventrális területre a P sejtek ingerületei tartanak. Az M sejtek inkább a mozgás és a gyors idői változások lekövetéséért felelősek, a P sejtek inkább a finom részletek, a szín, a textúra reprezentálásáért.
1.5. A színlátás
Jelen fejezetben szó lesz a színlátás fotoreceptorairól (pálcikák és csapok), a tárgyak színének észleléséről, az emberi színlátás opponens színelméletéről, és a konstanciákról a feldolgozás során.
1.5.1. A színlátás fotoreceptorai
Az emberi szem retináján négy különböző spektrális érzékenységű fotoreceptor található. A pálcikáknak csupán a fényintenzitás változásának detektálásában van szerepe. A három csaptípus azonban tökéletesen alkalmas arra, hogy megfelelő feldolgozás, összehasonlítás után képes legyen differenciálni a különböző frekvenciájú sugárzásokat, fényeket. A három csaptípusunk különböző spektrális érzékenységi görbével rendelkezik, amelyeket ezért hosszú, közép és rövid hullámhosszú csapoknak vagy kék, zöld és vörös csapoknak nevezünk. A három csapot különböző energiával érkező fotonok ingerlik. Az kék, zöld és vörös csapok érzékenységi maximumai rendre 440, 545 és 575 nm hullámhosszúságú, látható sugárzásnál vannak. A közepes és hosszú hullámhosszakra érzékeny csapok érzékenységi görbéi szinte egymásba simulnak. Ebből arra is következtethetnénk, hogy az ember evolúciója alatt kisebb változások, mutációk következtében az egykori két csaptípusból lassan differenciálódott egy harmadik, mert az jobban szolgálta a túlélést. A különböző típusú csapok bármilyen fotont is nyelnek el, minden esetben ugyanolyan válaszreakciót adnak. Ezt nevezzük az univariancia elvének. A színérzékelő receptoraink meglehetősen durva színdiszkriminációs képességgel rendelkeznek, ezért sok esetben két fizikailag eltérő színt pszichológiailag azonosnak ítélünk meg. Ezeket a színeket nevezzük metamereknek.
(Amennyiben az Olvasó több információt szeretne kapni az emberi látórendszerről, a megadott szakirodalmakon kívül az Optika TÁMOP tananyagunkat ajánljuk a figyelmébe.)
1.5.2. Mitől függ a tárgyak színe?
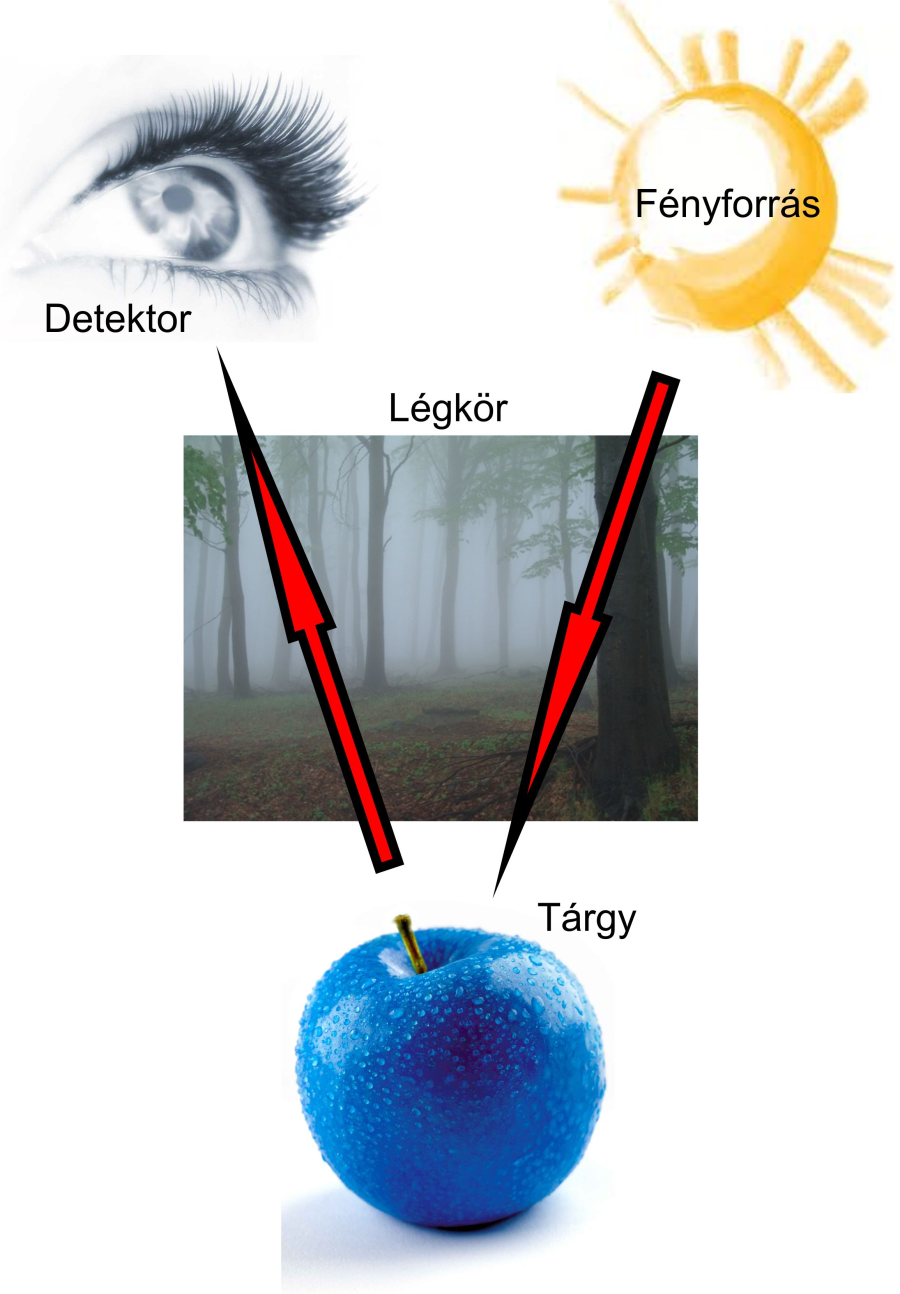
Tisztában kell lennünk azzal, hogy a színek észleléséhez sok lépcsőn keresztül vezet az út. Elsődleges a fényforrás, amely valamilyen spektrumú, összetett vagy monokromatikus fényt bocsát ki magából. Ez lehet természetes – Nap – vagy mesterséges fényforrás, mint például a halogén izzószálas vagy a LED fényforrás. Ezek a fényforrások adják azokat a frekvenciájú összetevőket, amelyek a tárgyakon visszaverődnek. A tárgyak felületének színe voltaképpen egy olyan görbével írható le, amely a beeső fényforrás spektrumából valamely hullámhosszakat visszaver, más hullámhosszakat elnyel. Ha fehér tárgyra fehér fénnyel világítunk, akkor a felület minden kibocsátott fényösszetevőt visszaver. Fekete felületről akkor beszélünk, ha a felület egyetlen hullámhosszon érkező fényösszetevőt sem ver vissza. A harmadik lépcsőfok a légkör, a közeg anyaga, amelyen keresztül a fény beérkezik. A világűrben például a rövid hullámhosszú fényösszetevők is haladhatnak, de a földi légkörben ezek jó része elnyelődik. A légkör tehát elnyeli a már szemükbe visszavert fény további alkotóelemeit, ezzel a visszavert fényösszetevők keveréke ismét módosul. A következő lépcsőfok az emberi szem, pontosabban a retina fotoreceptorainak érzékenysége. Szemünk is rendelkezik tehát egy érzékenységi görbével, amely megmutatja, hogy a különböző hullámhosszú fényösszetevőkre mekkora válaszreakciót várhatunk a receptorokról. A receptorokhoz érkező összetett fény színe pedig a receptorok reakcióinak összegzése után alakul ki, és még nem is beszéltünk a magasabb agyi folyamatokról. Látható tehát, hogy egy szín érzékelése mennyi mindentől függ, fizikai és pszichológiai téren egyaránt.
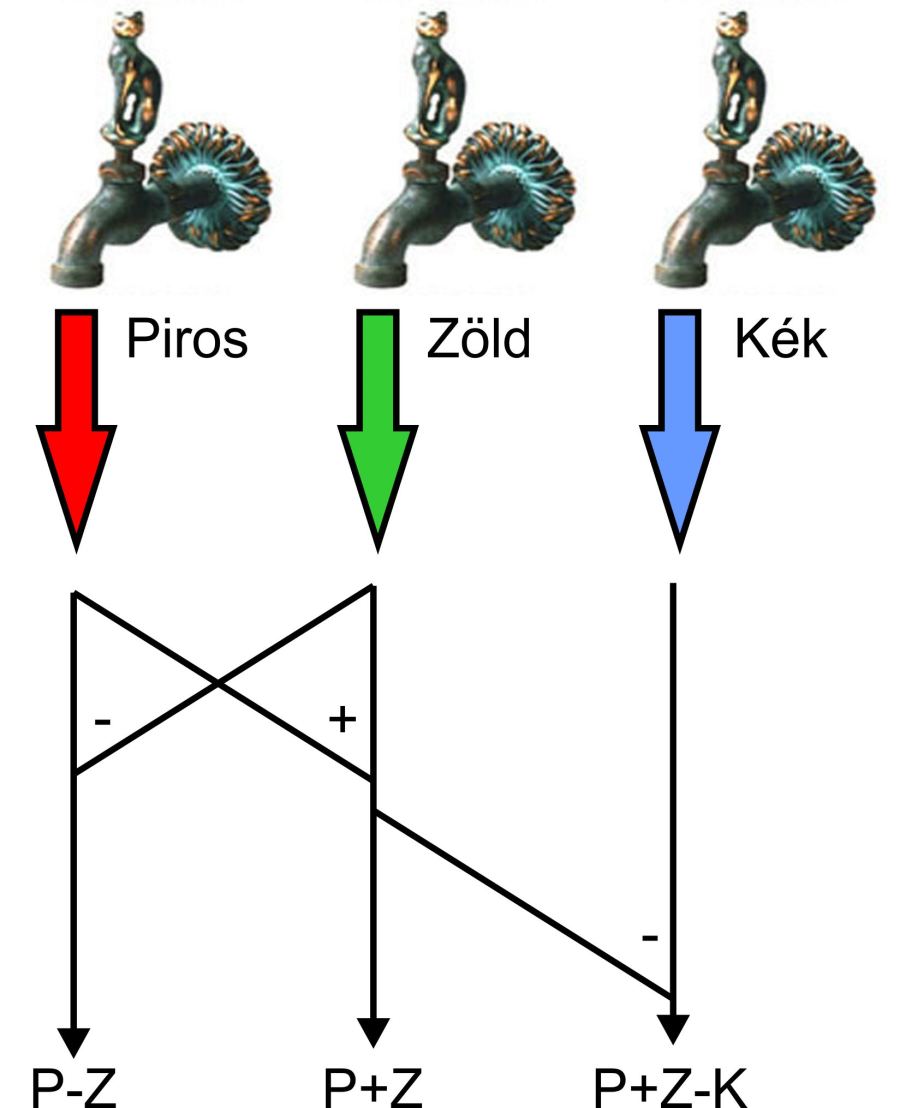
1.5.3. Az emberi színlátás opponens színelmélete
A színek kognitív feldolgozásának előszobája az ellenszín-elmélet – opponens – amelynek mai formáját Dorothea Jameson és Leo Hurvich dolgozták ki 1981-ben. A modell szerint látórendszerünk a csapok jeleiből különbségeket állít elő, és ezzel határozza meg az adott színárnyalatot. A piros és a zöld csapok összevetéséből előáll egy piros-zöld különbségjel. Ez alkalmas annak eldöntésére, hogy az adott árnyalat inkább piros, vagy inkább zöld-e, esetleg azonos mértékben mindkettő. Ez után következik az egyesített piros + zöld csatornajel összehasonlítása a kék csapok jelével. Ha a kék a nagyobb intenzitású, akkor a színárnyalat kékes, ha nem, akkor sárgás lesz, esetleg azonos mértékben mindkettő. Végül a piros + zöld csatornajel összegzett intenzitása adja meg az akromatikus világosságérzetet.
1.5.4. Konstanciák a feldolgozás során
Feldolgozó rendszerünk rendelkezik bizonyos képességekkel, amelyek elősegítik, hogy a látórendszer által érzékelt információt különböző körülmények között is azonosnak értékeljük. Például a sárga taxit minden esetben sárgának látjuk, noha változnak a megvilágítási viszonyok, vagy az írólap világosságát mindig ugyanolyannak látjuk, pedig időközben besötétedett. Az előbbit színkonstanciának (színállandóságnak), utóbbit pedig világosság-konstanciának (világosság-állandóságnak) nevezzük. Létezik egy normálási folyamat is feldolgozás során, amikor is a látórendszerbe érkező képen túlzottan érvényesülő színtartományokat képesek vagyunk egy szintre hozni. Például naplementekor a narancssárgás fények miatt azokon a hullámhossz tartományokon (narancssárgás tartományon) a feldolgozórendszer csökkenti az érzékenységet. Azt a tényt, hogy a feldolgozórendszerünk az érzékelt területet kisebb szegmentumokra bontja, mi sem mutatja be jobban, mint a szimultán kontrasztjelenség, amelynél látható, hogy egy felület színezetét nagyban befolyásolja, hogy a szín milyen környezetben, kontextusban szerepel a képen. Sötétebb kontextusban ugyanaz a világosság világosabbnak tűnik, mint világosabb közegben. A színkontraszt ellentétes jelensége a színasszimilációs hatással, amelynél az alapszínt körülvevő színes csíkok a saját színezetük irányába tolják el az alapszín színezetét. Ezek a jelenségek a színlátás során érvényesülő magasabb agyi folyamatok jelenlétére utalnak.

1.6. A háromdimenziós mélységészlelés
A mélységészlelés tárgyalása előtt nélkülözhetetlen néhány pszichológia alapfogalmat tisztáznunk. Az ember az őt körülvevő tárgyakat valamilyen formán észleli. Ennek a folyamatnak ez első lépése a tárgyak háttértől való elkülönítése, a perceptuális szegregációja. Ezeket a folyamatokat a V1 agyterület alapozza meg. Ez a terület felelős a körvonalak detektálásáért és a tárgyak háttértől való elkülönítéséért is. A tárgyak háttértől való kelkülönítésében nagy segítséget jelent a feldolgozórendszer számára, hogy a tárgy általában a háttér előtt helyezkedik el, a háttér a tárgy mögött is folytatódik, míg a körvonal általában a tárgyhoz tartozik és nem a háttérhez, valamint a tárgy jobban hasonlít egy már látott tárgyra, mint a háttér. Tehát a látott képen lévő tárgyak mérete, szimmetriája, orientációja és ismertsége nagyban hozzájárul ahhoz, hogy el tudjuk különíteni egymástól a hátteret és a tárgyat.
A mélységészlelés – távolságészlelés – tehát azon képességünk, amellyel a körülöttünk lévő világot három dimenzióban láthatjuk. Képesen vagyunk nagy pontossággal felmérni a tárgyak távolságát, megfogni dolgokat, illetve mozogni a térben. A mélységészlelés képességét már a háromhónapos csecsemőknél ki lehet mutatni, ami azt jelenti, hogy ez biológiailag is igen fontos képességünk.
Jelen fejezetben szó lesz a Gestalt-elméletről, a monokuláris és binokuláris jelzőmozzanatokról valamint a binokuláris diszparitásról.
1.6.1. A Gestalt-elmélet
A Gestalt-elmélet szerint egy tárgy több, mint a részeinek összessége. A tárgyak felismerésében nagy szerepe van az alábbi, úgynevezett Gestalt-szabályoknak. Az adott mintázatot úgy látjuk, ahogy az a lehető legegyszerűbb. Az egymáshoz hasonlatos mintarészeket egymással csoportosítva látjuk. Az egy ívre illeszkedő részeket egy folytonos alakként értékeljük. Az egymáshoz közelebbi tárgyakat egyként kezeljük. Az együtt mozgó tárgyakat egyként kezeljük. Ha az előzőekben ismertük a tárgyak, akkor könnyebb a későbbi felismerés és a részek csoportosítása, az egész előállítása érdekében.
1.6.2. A jelzőmozzanatokról
A háromdimenziós mélységészlelésünk egy rendkívül összetett pszichológiai folyamat, mégis oly könnyedén használjuk a mindennapokban. Működtetése nem igényel tudatos erőfeszítést, teljesen automatikus, és mentálisan nem növeli a megterhelést. A tér és mélységészlelés azon folyamatok egyike, amely talán a leginkább rávilágít arra, hogy látásunkban csupán kis részt vállal maga a látószerv és hatalmas részt az agy. A térérzékelésben két fő jelzőmozzanat-típust különböztethetünk meg. Az egy szemmel is működő monokuláris és a két szemmel használható binokuláris jelzőmozzanatokat. Mint már utaltunk rá, az állatvilágban a növényevőknek általában a fejük két oldalán helyezkedik el a szemük, hiszen létfenntartásukhoz sokkalta fontosabb a nagy látótér, mint a mélységérzékelés. A ragadozóknak azonban fontosabb, hogy meg tudják becsülni prédájuk távolságát, így érthető, hogy az evolúció folytán a fejük elülső részére kerültek a szemek egymással egy síkban, hogy így a lehető legnagyobb binokuláris teret hozzák létre. Persze látnunk kell azt is, hogy a binokuláris látásunk csupán korlátozott távolságon belül használható hatékonyan, ezen kívül nem jobb, mint az egyszemes látás.
Az álltavilágban sok módját találhatjuk annak, hogy bizonyos fajok milyen módon érzékelik környezetüket. Gondolhatunk itt például a denevérekre, vagy a teljesen vak barlangi rákokra. De kibővíthetjük látókörünket azzal is, hogy az ember miként képes az őt körülvevő tér érzékelésére szemek nélkül - a karjait, a hangját, a kopogtatást és egyéb eszközöket igénybe véve. Számunkra azonban jelen téma kapcsán csupán a látás a fontos. Meg kell ismernünk mindazokat a folyamatokat, amelyek hozzásegítenek bennünket a térérzékeléshez.
1.6.3. Monokuláris jelzőmozzanatok
Az alábbiakban áttekintjük a monokuláris jelzőmozzanatokat, megnézzük, hogy miből tudunk következtetni a tárgyak távolságára, ha csak egy szemmel nézzük azokat. Ilyenek a tárgyak retinális nagysága és relatív retinakép mérete, a horizonthatás, a takarási hatás, a perspektíva, a textúra, az elkékülési hatás, a légtávlat, a fény-árnyék játék, az akkomodáció és a mozgási parallaxis.
A retinális nagyság: Elsődleges jelzőmozzanat, hogy a térben elhelyezkedő tárgy mekkora területet foglal el a retinán. Ebből a feldolgozórendszerünk meg tudja becsülni az észlelt tárgy távolságát, amennyiben rendelkezik priori (előzetes) információval annak tulajdonságait illetően. Ha például egy kisautót látunk magunk előtt az asztalon, akkor felismerjük, hogy kisautó és nem gondoljuk azt, hogy egy valódi gépkocsi a távolban.

A horizont hatás: A tárgyak távolságának becslésében az is segítség, hogy a látómezőnk horizontja fölött vagy alatt helyezkedik el. Minél feljebb van ugyanis, annál távolabbinak látjuk a tárgyat, és minél lejjebb van, annál közelebbinek.

A takarásihatás: Ha egy tárgy eltakarja egy másik tárgy bizonyos részeit, akkor arra következtetünk, hogy közelebb van hozzánk. Ez a takarás jelensége.

Perspektíva: Szintén tanult és mélyen rögzült jelenség, hogy a párhuzamosnak gondolt élek a távolban összetartanak. Ez a lineáris perspektíva, amellyel becslést tudunk adni a tárgyak egymáshoz képesti elhelyezkedésére.

Textúra: A textúrát személve észrevehetjük, hogy a közelebbi textúrákat jóval részletgazdagabbaknak találjuk, mint a távolabbi textúrákat. A részletgazdagsággal együtt természetesen a textúra elemeinek mérete is a távolsággal fordítottan arányos. Ha például egy macskaköves utat szemlélünk, láthatjuk, hogy a távolabbi kövek kisebbek és kevésbé részletgazdagok.

Az elkékülésihatás: A Nap összetett fénye a Föld légkörén áthaladva szóródik. A szóródás a kék tartományokban magasabb, így a nappali légkört kéknek látjuk. Ha egy távoli tárgy és közöttünk nagy légtömeg van, a tárgy képe elkékül. Ennek a kékülésnek a mértéke arányos a tárgy távolságával. Ezt az úgynevezett elkékülési hatást használhatjuk a távoli tárgyak, például hegyek távolságának becslésére.
Légtávlat: A távoli tárgyak nem csupán kékülnek a közbenső légtömeg miatt, hanem a kontrasztjuk is romlik, éleik elmosódnak, és színeik tompulnak. Az elkékülési hatással együtt ezt a jelenséget légtávlatnak nevezzük.

A fény-árnyék játék: További segítség lehet egy tárgy térbeli elhelyezkedésének valamint kiterjedésének becslésére, a tárgyon feltűnő árnyék és fény viszonya, határvonala, illetve annak tranziense.

A relatív retinakép mérete: Ha látunk két tárgyak, amelyről nincsenek priori információink, illetve azonos méretűnek gondolnánk azokat, akkor a két tárgy közül azt látjuk nagyobbnak, amelyik mérete a retinánkon nagyobb.
Akkomodáció: Egy kevésbé kognitív jelzőmozzanat lehet a szemlencse akkomodációja. Ez a folyamat akkor következik be, amikor szemünkkel élesre szeretnék állni egy tárgyra valahol a térben. Ebben az esetben a szemlencse feszítőizmai úgy ernyednek el és feszülnek meg, hogy az egyik legtökéletesebb szabályozókört alkalmazva úgy állítják be a szemlencse görbületi sugarát, hogy a tárgy képe pontosan a retinára képeződjön le. Ez hasznos információ lehet a feldolgozórendszerünk számára a közelebbi – körülbelül 2 méterig bezárólag – tárgyak távolságának becslésére.
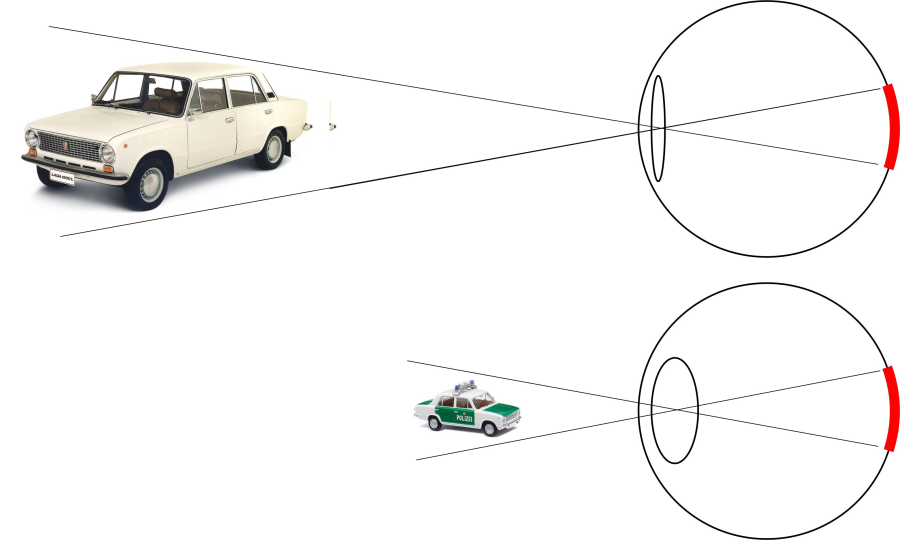
A mozgási parallaxis: Az azonos sebességgel mozgó tárgyak közül azt érzékeljük közelebbinek, amely gyorsabban halad át a látómezőnkön – a másikhoz képest. Ekkor a retinánkon egyfajta szögsebességet mérhetnénk, hiszen a retina gömbfelülethez hasonló alakú. Gondolhatunk például arra, hogy utazás közben a vonatból kitekintve a közeli fák nagyobb szögsebességgel haladnak át a retinán, mint a távoli házak. Ezt a jelenséget mozgási parallaxisnak nevezzük.

1.6.4. Binokuláris jelzőmozzanatok
Az egyik legegyszerűbb binokuláris jelzőmozzanat a konvergencia. Amikor a két szemünkkel egy 6-10 méter távolságon belüli tárgyra nézünk, látórendszerünk a két szem tengelyét igyekszik összetartóan a tárgyra fordítani. Ennek a konvergenciának a foka fontos jelzőmozzanat a tárgy távolságának becslésére. Amennyiben a két szemtengely által bezárt szög kisebb, a tárgy távolabb van, amennyiben a szög nagyobb, a tárgy közelebb van.
A másik jelzőmozzanat a binokuláris parallaxis. A parallaxis eredendően a testek egymáshoz viszonyított helyzetének változása eltérő irányokból nézve. Tegyük fel, hogy a két szemünkkel egy tárgyra nézünk a térben! Emellett megjelenik egy másik tárgy is, amely az előzőhöz viszonyítva más-más helyen bukkan fel, attól függően, hogy melyik szemünkkel nézzük. Ha a két tárgy azonos távolságban van, akkor a két szemünkkel ugyanazt az elhelyezkedést látjuk. Ha a két tárgy különböző távolságban található, akkor az egyikre nézve, és a két szemet egyenként kinyitva azt tapasztaljuk, hogy a tárgyak távolsága változik a szemeink által látott képen. Ennek a vízszintes távolságváltozásnak, az úgynevezett parallaxisnak annál nagyobb a mértéke, minél nagyobb volt a két tárgy közötti különbség. Valós helyzetben persze minden esetben valamelyik tárgyra akkomodál a szemünk, ezért a másik tárgyat nem láthatjuk tisztán. Minél nagyobb volt a távolság, annál homályosabban látjuk azt. A háromdimenziós mozifilmeken ezt nem tapasztaljuk, hiszen szemlencsénk ilyenkor minden esetben a mozivászonra akkomodál.
Binokuláris jelzőmozzanat továbbá a tárgyak nézete is. Amikor két szemünkkel nézzük a közeli tárgyakat, akkor szemeink különböző nézetekből látják azokat, és ezért a retinánkra érkező kép nem csupán parallaxist tartalmaz, hanem a tárgyak más-más oldalának képét is. Egyik szemünkkel láthatunk olyan részletet a tárgyról, amit a másik szemünkkel nem, és fordítva. Ez a két kép közötti apró különbség is hozzájárulhat a tárgyak távolságának észleléséhez.
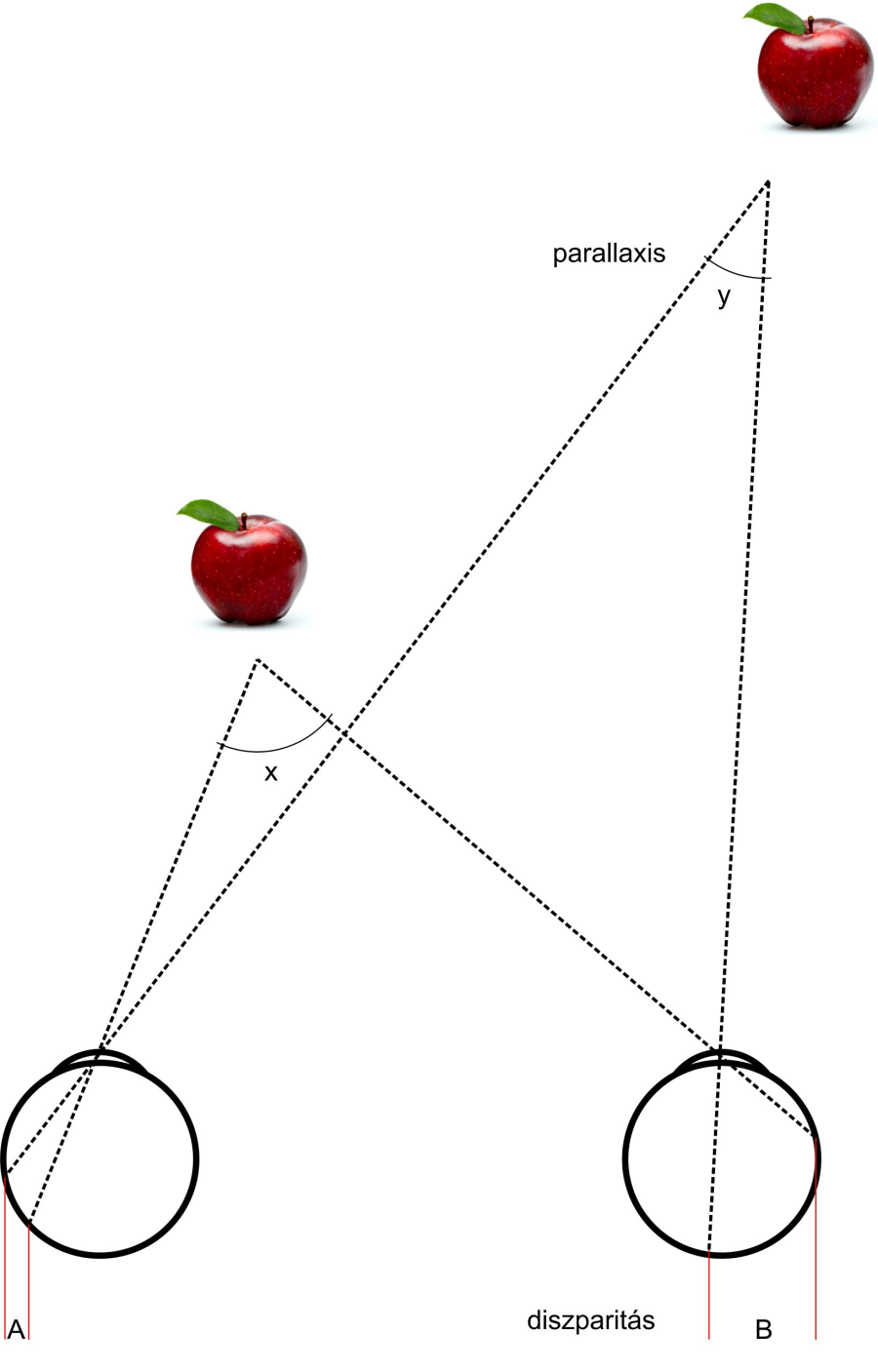
1.6.5. A binokuláris diszparitás
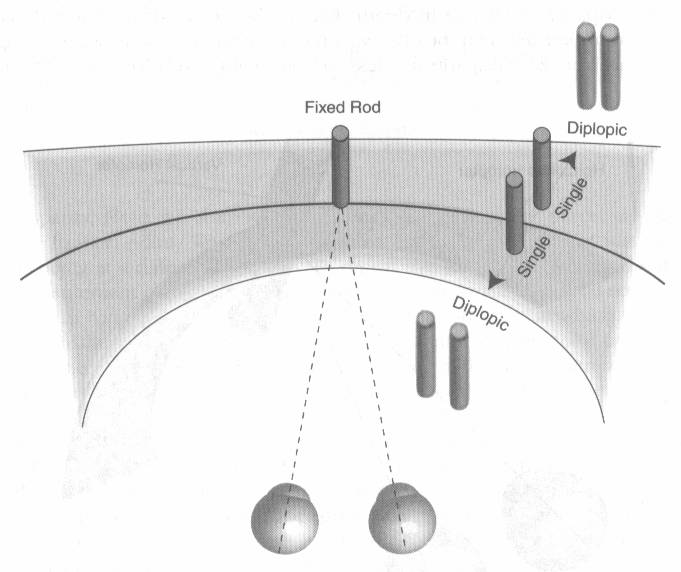
A diszparitás jelentése eltérés. Eltérés a két szem retinális képe között, pontosabban a két képen lévő tárgyak távolsága között. Úgy gondolhatnánk, hogy a teret teljes egészében térben látjuk, valójában azonban a teljes tér csak egy csekély szeletét látjuk valós sztereolátással. Azt a jelenséget, hogy két szemünk retináján két kissé különböző kép keletkezik, binokuláris diszparitásnak nevezzük. Horopternek hívjuk azt a térbeli felületet, amelynek minden pontját azonos szögből látjuk a két szemünkkel. Ha rögzítünk egy pontot a térben láthatjuk, hogy a tárgytól nagyobb vagy kisebb mélységben lévő egyéb tárgyakból kettőt látunk. A fixált tárgy körül van egy mező, amelyen belül működhet a valódi sztereolátásunk, azon kívül a képek szétesnek, és kettős látás jön létre. Ezt a mezőt sztereomélységnek hívjuk. Panum 1858-ban igazolta, hogy idegrendszerünkben létezik egy, az előbbivel összefüggésbe hozható mező. Ez azért lehetséges, mert az idegrendszerünkben létrejövő szerveződés retinotopikus, azaz a retina szomszédos receptorainak ingerületei az agy szomszédos neuroncsoportjaihoz kapcsolódnak. A sztereolátásért felelős agyi terület mikroszkóp alatt is láthatóan csíkozott, a két szem által alkotott képek ugyanis az egyik és a másik szem által küldött képrészleteket felváltva tartalmazzák.
Julesz Béla 1960-ban felfedezte, hogy a sztereolátást nem előzi meg az tárgyak azonosítása, mint ahogy azt addig hitték. A térbeli mélységet a retinális képeken lévő tárgyak felismerése nélkül is képesek vagyunk érzékelni. Julesz ezt a felfedezést a róla elnevezett random-dot (véletlen pont) sztereopárokkal bizonyította be. A Julesz-féle sztereopár két képet tartalmaz a két szem számára, amelyet szeparáltan kell bemutatni. Az egyik képen véletlen pontokat látunk, a másik képen pedig az előző kép kicsit módosított változatát. A módosítás lényege, hogy a kívánt alakzat pontjait a diszparitás mértékével eltoljuk balra vagy jobbra, és az üresen maradt képpontokat véletlenszerűen feltöltjük. Ekkor a személőnek azonnal előbukkan a kívánt alakzat, az előzetes felismerés nélkül. A kép nagyon stabil, pásztázó szemmozgásoknál is fennmarad.

1.7. Az emberi mélységészlelés hibái
Eddig áttekintettük a háromdimenziós megjelenítési technikák emberi tényezőit. Azonban nem tettünk említést arról, hogy az emberi mélységészlelés nem minden esetben működik. Vannak olyan tényezők, amelyeknek mindenképpen jól kell működniük, például a háromdimenziós filmek megfelelő észleléséhez. A különböző látáshibák, amelyek a mélységészlelést befolyásolják, nem csupán a háromdimenziós élményt veszik el az embertől, hanem a vetítés után további kellemetlen perceket, órákat is hoznak azzal, hogy émelygést, fejfájást és hányingert okoznak. Egy átlagos háromdimenziós film vetítésénél mindig akad egy-két ember a nézőtéren, aki nem látta három dimenzióban a filmet. Ez nem is meglepő, hiszen egy amerikai felmérés szerint a 18 és 38 év között lakosság 56%-a szenved valamilyen mélységészlelést érintő tünettől – a teljes lakosság hozzávetőlegesen 5-10%-át érintheti. Mi lehet az a tényező, amely nem engedi, hogy a mélységészlelési zavarokkal küzdő emberek három dimenzióban lássák a háromdimenziós filmeket? Az ő látórendszerük valamilyen oknál fogva – lehet az látószervi, de akár központi idegrendszeri zavar is – nem képes a két szemnek küldött képeket „összeolvasztani”.
Az egyik probléma, amely korlátozhatja a háromdimenziós élményt az amblyopia – lusta szem. Ebben az esetben az egyik szem sokkalta rosszabb képet érzékel – például 2-3 dioptriával rosszabb a másiknál. Ilyenkor a látórendszer nem képes összerakni a két szem képét, vagy súlyosabb esetben „szoftveresen lekapcsolja” a rosszabbik szemet, amikor mindkét szem nyitva van, és csak a jobbik szem képéből dolgozik. Az ilyen látáshibát a tünettel rendelkezőnek nehéz észrevenni, mert már megszokta.
A jelenséget kiválóan demonstrálja a binokuláris versengés jelensége, ahol a két szemnek teljesen különböző képeket adunk, és a két szem képe gyakorlatilag „verseng” a feldolgozásért. Egyszer az egyik jelenik meg, egyszer a másik.
Egy másik ok lehet a háromdimenziós élmény hiányára a kancsalság. Ebben az esetben a két szem nem azonos irányba áll, amikor egy tárgyra fokuszál. Ilyenkor kettős látás jelentkezhet, és a háromdimenziós élmény teljes mértékben hiányozhat. További probléma lehet, ha a szemek képtelenek megfelelően egymás felé fordulni – konvergencia-elégtelenség.
Az efféle látáshibák nagy részét lehet kezelni, azonban ennek előfeltétele, hogy észrevegyék. Ebben viszont igen nagy segítséget nyújtanak a háromdimenziós mozik, ahol az ilyen aberrációkra rövid úton fény derül.
Amikor egy háromdimenziós filmet nézünk, szemünkkel a vetítővászonra – illetve a képernyőre, vagy a nyomtatott médiumra – akkomodálunk, viszont szemeink vergeciája ezzel nincs szinkronban, mint valós körülmények között. Tehát a szemlencsénk egy adott távolságra van „beállítva, míg a szemeink széttartása a háromdimenziós tartamom függvényében más-más távolságokra van hangolva. Ennek pedig sok esetben rosszullét, fejfájás, szédülés és hányinger lehet a következménye, főleg a „gyakorlatlan” szemű embereknél.

1.8. Összefoglalás
Az érzékelés az a folyamat, amely során a külvilágból érkező információk valamely érzékszervünkön keresztül elérhetővé válnak a feldolgozórendszerünk számára. Az észlelési folyamat magából az érzékelésből és a hozzá kapcsolódó kognitív feldolgozásból áll.
Az emberi szem a 380 és 780 nm közötti hullámhossztartományt képes érzékelni. Az emberi szem a fej elülső oldalán helyezkedik el, egymással egy síkban és egymástól 63 mm-es távolságban. Látómezőnk közel 90 fokban sztereolátást tesz lehetővé. Szemünk mozgását a szem körül elhelyezkedő három pár extraokuláris izom teszi lehetővé. A konjunktív szemmozgás során két szemünk egy irányba mozog, vergens szemmozgás során szemünk a közeledő, illetve távolodó tárgyakra néz. Szemünk kétféle szemmozgást képes kivitelezni. Az egyik egy szakaszos, szaggatott, gyors szemmozgás, a másik egy folyamatos mozgás. A szakaszos szemmozgás ugrásait szakkádoknak, a megállásait fixációnak nevezzük. A gyors szemmozgások alatt nincs információfelvétel. A követő szemmozgáskor a mozgás sebessége és iránya folyamatosan változik. Az emberi szem három koncentrikus rétegből, két kamrából, a szivárványhártyából, a pupillából és a lencséből áll. A fotoreceptorok a retinán helyezkednek el. A retinán az éleslátás helye az úgynevezett fovea. Ahol a gyűjtősejtek idegrostjai elhagyják a retinát az agy felé, nincsenek fotoreceptorok. A retinán lévő fotoreceptorok száma 130 millió. A receptorok két fő típusa a csapok és pálcikák. A pálcikák több fotont képesen elnyelni több irányból, egy ganglionsejthez több pálcika idegrostja fut be. A csapok három altípusát különböztetjük meg. Léteznek rövid, közép és hosszú hullámhosszakra érzékeny csapok. Az éleslátás helyén szinte csak csapok vannak, a periférián szinte csak pálcikák. A fotoreceptorok neuronális jelei a gyűjtősejteken keresztül a retinális ganglionsejtekhez jutnak. A ganglionsejtek száma körülbelül 1 millió. A ganglionsejtekhez kapcsolódó fotoreceptorok egy kvázi kör alakú, úgynevezett receptív mezőt alkotnak, amelynek a közepén lévő receptorokat ingerelve növelhető, a perifériát ingerelve csökkenthető a ganglionsejt reakciója. A retina fotoreceptorai receptív mezőkbe tömörülve térbeli integrációt végeznek. A retina és a látókéreg között a látópálya teremt kapcsolatot. A retinális ganglionsejtek axonjai alkotják tehát a látóideget. A V1 látórendszer egy úgynevezett retiotopikus reprezentációval rendelkezik. A ganglionsejteknek van egy kiterjedtebb dendritfával és egy kisebb dendritfával rendelkező típusa. A nagyobb receptív mezővel rendelkező ganglionsejteket M sejteknek, a kisebb receptív mezővel rendelkezőket pedig P sejteknek hívjuk. Az M sejtek a CGL terület magnocelluláris rétegéhez, a P sejtek a CGL parvocelluláris rétegéhez kapcsolódnak. Mivel az M sejtekhez több receptor tartozik, azért a térbeli felbontóképességük rosszabb, mint a P sejteknek, viszont az M sejtek időbeli felbontóképessége sokkal jobb, mint a P sejteké, mert aktivitásuk kevesebb ideig marad fönn. Továbbá, az M sejtek csak világosságot kódolnak, míg a P sejtek színeket is. A V1 területről az információk egy része az agy felső részébe, a parientális lebenybe tart, a dorzális területre, egy másik részük pedig az agy alsó részébe, a temporális lebeny ventrális területére. A dorzális területre az M sejtek tartanak, a ventrális területre pedig a P sejtek. Az M sejtek inkább a mozgás és a gyors időbeli változások lekövetéséért felelősek, a P sejtek inkább a finom részletek, a szín, a textúra reprezentálásáért. Az emberi szem retináján négy különböző spektrális érzékenységű fotoreceptor található. A három csaptípusunk különböző spektrális érzékenységi görbével rendelkezik, amelyeket ezért hosszú, közép és rövid hullámhosszú csapoknak vagy kék, zöld és vörös csapoknak nevezünk. Látórendszerünk a csapok jeleiből különbségeket állít elő, és ezzel határozza meg az adott színárnyalatot. Feldolgozórendszerünk rendelkezik bizonyos képességekkel, amelyek elősegítik, hogy a látórendszer által érzékelt információt különböző körülmények között is azonosnak értékeljük.
A térérzékelésben két fő jelzőmozzanat típust különböztethetünk meg. Az egy szemmel is működő monokuláris és a két szemmel használható binokuláris jelzőmozzanatokat. A binokuláris látásunk csupán korlátozott távolságon belül használható hatékonyan, ezek kívül nem jobb, mint az egyszemes látás.
Elsődleges monokuláris jelzőmozzanat, hogy a térben elhelyezkedő tárgy mekkora területet foglal el a retinán. A tárgyak távolságának becslésében az is segítség, hogy a látómezőnk horizontja fölött vagy alatt helyezkedik el. Ha egy tárgy eltakarja egy másik tárgy bizonyos részeit, akkor arra következtetünk, hogy az közelebb van hozzánk. Szintén tanult, és mélyen rögzült jelenség, hogy a párhuzamosnak gondolt élek a távolban összetartanak. A textúrát személve észrevehetjük, hogy a közelebbi textúrákat jóval részletgazdagabbaknak találjuk, mint a távolabbiakat. A Nap összetett fénye a Föld légkörén áthaladva szóródik. A szóródás a kék tartományokban magasabb, így a nappali légkört kéknek látjuk. Ha egy távoli tárgy és közöttünk nagy légtömeg van, a tárgy képe elkékül. Ennek a kékülésnek a mértéke arányos a tárgy távolságával. A távoli tárgyak nem csupán kékülnek a közbenső légtömeg miatt, hanem a kontrasztjuk is romlik, éleik elmosódnak és színeik tompulnak. További segítség lehet egy tárgy térbeli elhelyezkedésének és kiterjedésének becslésére, a tárgyon feltűnő árnyék és fény viszonya, határvonala, illetve annak tranziense is. Ha látunk két tárgyat, amelyről nincsenek priori információink, illetve azonos méretűnek gondolnánk azokat, akkor a két tárgy közül azt látjuk nagyobbnak, amelyik mérete a retinánkon nagyobb. Egy kevésbé kognitív jelzőmozzanat lehet a szemlencse akkomodációja. Az azonos sebességgel mozgó tárgyak közül azt érzékeljük közelebbinek, amely gyorsabban halad át a látómezőnkön.
Az egyik legegyszerűbb binokuláris jelzőmozzanat a konvergencia. Amikor a két szemünkkel egy 6-10 méter távolságon belüli tárgyra nézünk, látórendszerünk a két szem tengelyét igyekszik összetartóan a tárgyra fordítani. Ennek a konvergenciának a foka fontos jelzőmozzanat a tárgy távolságának becslésére. A másik jelzőmozzanat a binokuláris parallaxis. Ha két szemünkkel egy tárgyra nézünk a térben, és megjelenik egy másik tárgy is, amely az előzőhöz viszonyítva más-más helyen bukkan fel, attól függően, hogy melyik szemünkkel nézzük. Ha a két tárgy azonos távolságban található, akkor a két szemünkkel ugyanazt az elhelyezkedést látjuk. Ha a két tárgy különböző távolságban van, akkor az egyikre nézve, és a két szemet egyenként kinyitva azt tapasztaljuk, hogy a tárgyak távolsága változik a szemeink által látott képen. Ennek, a vízszinten távolságváltozásnak, az úgynevezett parallaxisnak annál nagyobb a mértéke, minél nagyobb volt a két tárgy közötti különbség. Binokuláris jelzőmozzanat továbbá a tárgyak nézete is. Amikor két szemünkkel nézzük a közeli tárgyakat, akkor szemeink különböző nézetekből látják a tárgyakat, és ezért a retinánkra érkező kép nem csupán parallaxist tartalmaz, hanem a tárgyak más-más oldalának képét is.
A teljes tér csak egy csekély szeletét látjuk igazi sztereolátással. Azt a jelenséget, hogy két szemünk retináján két, kissé különböző kép keletkezik, binokuláris diszparitásnak nevezzük. Horopternek hívjuk azt a térbeli felületet, amely minden pontját azonos szögből látjuk a két szemünkkel. A fixált tárgy körül van egy mező, amelyen belül valódi sztereolátásunk működhet, azon kívül a képek szétesnek, és kettős látás jön létre.
1.9. A felkészülést segítő kérdések
-
Mi a különbség az érzékelés és az észlelés között?
-
Mi az a szignál transzdukció?
-
Mik azok a receptorok?
-
Mit jelent az adekvát inger kifejezés?
-
Mi az a percepció?
-
Igaz-e, hogy észlelés történhet érzékelés nélkül?
-
Az emberi szem milyen intervallumban képes érzékelni az elektromágneses spektrumot?
-
Igaz-e, hogy a nagyfrekvenciájú fény, nagy hullámhossz értékű is?
-
Hozzávetőlegesen milyen szögben van térlátásunk?
-
Milyen izmok teszik lehetővé a szem mozgását?
-
Mi az a konjunktív szemmozgás?
-
Mi az a vergens szemmozgás?
-
Hogy nevezzük a közvetlenül előttünk lévő tárgya történő fókuszálást?
-
Mikor használunk szakaszos, és mikor folytonos szemmozgást (példákkal)?
-
Mi az a szakkádikus elnyomás?
-
Igaz-e, hogy a fixációk alatt nincsen információfelvétel?
-
Mi az a mikroszakkádikus mozgás?
-
Mi történik, ha a mikroszakkádikus mozgást kiiktatjuk?
-
Igaz-e, hogy a cornea az ínhártya része?
-
A szaruhártya vagy a szemlencse nagyobb törőértékű?
-
Igaz-e, hogy a csarnokvíz nem cserélődik?
-
Igaz-e, hogy az üvegtestfolyadék nem cserélődik?
-
Írja le a szemlencse görbületi sugarát változtató folyamatot!
-
Hol helyezkednek el a fotoreceptorok?
-
A fotoreceptoroktól hová tart az idegi jel?
-
Igaz-e, hogy a retina fény felé néző oldalán helyezkednek el a ganglionsejtek?
-
Igaz-e, hogy a fotoreceptorok sűrűsége nagyobb azon a helyen, ahol az idegek elhagyják a retinát?
-
Hozzávetőlegesen mennyi fotoreceptor van egy emberi szemben?
-
Igaz-e, hogy a fotoreceptorok sűrűsége a foveán a legnagyobb?
-
Igaz-e, hogy a színtévesztés leggyakoribb típusa a vörös-kék színtévesztés?
-
Igaz-e, hogy a retinán található fotoreceptorok között sokkal több csap van?
-
Igaz-e, hogy a sárgafolton sokkal több csap van, mint a retina más részein?
-
Milyen típusú csapok vannak, és milyen frekvenciatartományokban érzékenyek?
-
Igaz-e, hogy a kékes fényben a receptorok nagy száma miatt jobban látunk?
-
A retina mely területein érzékelünk jobban mozgásokat és fényintenzitás-változást?
-
A retina mely területein érzékelünk jobban színeket?
-
Átlagosan hány retinális ganglionsejtünk van?
-
Hogyan korrigálják az emmetropiás szem aberrációját?
-
Hogyan korrigálják az hiperópiás szem aberrációját?
-
Hogyan korrigálják az miópiás szem aberrációját?
-
Mi az a receptív mező?
-
Receptív mező tekintetében milyen retinális ganglion típusokat ismer?
-
Igaz-e, hogy a szemekbe balról érkező fénysugarak a bal agyféltekébe tartanak?
-
Milyen szerepe van annak, hogy látási információk tartanak a vegetatív idegrendszerbe?
-
Mi az a retiotopikus reprezentáció?
-
Mit nevezünk agykérgi nagyításnak?
-
Mik azok a P sejtek, és milyen tulajdonságokkal rendelkeznek?
-
Mik azok az M sejtek, és milyen tulajdonságokkal rendelkeznek?
-
Igaz-e, hogy a P sejtek ingerületei a dorzális területre tartanak?
-
Mit nevezünk az univariancia elvének?
-
Mik azok a metamerek?
-
Mi határozza meg, hogy milyen színben látunk egy tárgyat?
-
Fejtse ki az opponens színelméletet!
-
Mi az a konstancia?
-
Milyen konstancia típusokat ismer?
-
Mi az a perceptuális szegregáció, melyik agyterület felelős érte?
-
Mi az a mélységészlelés?
-
Milyen Gestalt-szabályokat ismer?
-
Miket nevezünk jelzőmozzanatoknak?
-
Mit nevezünk monokuláris jelzőmozzanatnak?
-
Mit nevezünk binokuláris jelzőmozzanatnak?
-
Milyen monokuláris jelzőmozzanatokat ismer?
-
Milyen binokuláris jelzőmozzanatokat ismer?
-
Melyik jelzőmozzanatnak jut főszerep a háromdimenziós mozikban?
-
A mozgási parallaxis egy binokuláris jelzőmozzanat?
-
Mi a binokuláris parallaxis?
-
Mi az akkomodáció?
-
Mit jelent a szemek konvergenciája?
-
Mi a binokuláris diszparitás?
-
Mit nevezünk horopternek?
-
Milyen háromdimenziós megjelenítés köthető Julesz Béla nevéhez?
-
Mi a random-dot sztereopár?
-
Mely esetekben fordulhat elő, hogy valaki nem képes a háromdimenziós tartalmak észlelésére?
-
Látórendszeri aberrációval nem rendelkező személyeknél mi lehet az oka a háromdimenziós tartalmak szemlélése közbeni rosszullétnek, szédülésnek?
1.10. A fejezet fontosabb kifejezései
|
adekvát inger agykérgi nagyítás akkomodáció amakrin sejtek amblyopia axon binokuláris diszparitás binokuláris jelzőmozzanat binokuláris parallaxis bipoláris sejtek caelsma opticum ciliáris izmok cornea corpus geniculatum laterale csapok csarnokvíz dendritfa depolarizálódás dorzális terület elülső szemkamra emmetropiás szem érhártya érzékelés észlelés extraokuláris izom fixáció folytonos szemmozgás foton fotoreceptor fovea ganglionsejtek Gestalt-szabályok gyűjtősejtek Hermann-rács hiperópiás szem hipotalamusz horopter horozontális sejtek idői felbontóképesség ínhártya jelátviteli kaszkád jelzőmozzanat kamrai folyadék konjunktív szemmozgás konstancia konvergens szemmozgás lencsefüggesztő rostok |
M sejtek macula magnocelluláris réteg mélységészlelés metamerek mikroszakkádikus mozgás miópiás szem monokuláris jelzőmozzanat mozgási parallaxis nátriumcsatornák okcipitális lebeny opponens színelmélet opszin P sejtek pálcikák parientális lebeny parvocelluláris réteg percepció perceptuális szegregáció perspektíva pupilla random-dot sztereopár receptív mező receptor retina retinal retinális kép retiotopikus rodopszin sárgafolt szakaszos szemmozgás szakkádikus elnyomás szakkádok szaruhártya szemlencse szignál transzdukció színdiszkriminációs képesség szivárványhártya temporális lebeny téri felbontóképesség téri integráció univariancia üvegtest V1 terület vakfolt ventrális terület vergens szemmozgás |
Felhasznált irodalmak
[1.1.] Pszichológia. Osiris Kiadó. Budapest . 2003.
[1.2.] A lélektan története. Osiris Kiadó. Budapest . 2010.
[1.3.] Affektív pszichológia. Medicina Könyvkiadó. Budapest . 2013.
[1.4.] Személyiségpszichológia. Osiris Kiadó. Budapest . 2006.
[1.5.] Fejlődéslélektan. Osiris Kiadó. Budapest . 2003.
[1.6.] Általános pszichológia 1. Osiris Kiadó. Budapest . 2007.
[1.7.] Szociálpszichológia. Osiris Kiadó. Budapest . 2004.